見逃してしまったアニメやドラマをもう一度

見たいと思っていたドラマやアニメを見逃したことはありませんか?
録画予約を忘れて見逃した作品はありませんか?
思い出の作品を見たいと思ってレンタルショップに行き、置いてないことにがっかりした経験はありませんか?
そんなとき、家にいながらすぐに見たい作品を見ることができるのが、動画配信サービス、通称VOD(ビデオ・オン・デマンド)です。

皆様お馴染みのAmazonプライムビデオやHulu(フールー)、近年人気を爆発させているNetflix(ネットフリックス)などサービスは数多くあり、配信されている作品も様々。
そんな中からどの動画配信サービスを選んだらいいのか、迷っている方も多いことと思います。
当サイトでは、そんな皆様のために、各配信サイトの利用料金、配信タイトルの傾向、特典など様々な角度から比較を行い、おすすめの動画配信サービスをご紹介いたします。
また、各配信サイトの最新の配信情報も随時更新してまいりますので、是非ともご活用ください。
おすすめ動画配信サービス比較表
まずは当サイトおすすめの『』、『Amazonプライムビデオ』、『hulu』の3サービスについて、簡単な比較表をご用意いたしました。
 |
 |
 |
|
|---|---|---|---|
| 作品数 | 80,000以上 | 非公開 | 50,000以上 |
| 最大画質 | HD | 4K | HD |
| オリジナル コンテンツ |
無 | 有 | 無 |
| 対応 デバイス |
Windows Mac iOS Android 他 |
Windows Mac iOS Android 他 |
Windows Mac iOS Android 他 |
| 月額料金 | 1,990円 ※税抜 |
400円 ※税込 |
933円 ※税抜 |
| 無料期間 | 31日間 | 30日間 | 2週間 |
ご覧のように、金額面だけを見るならば圧倒的にプライムビデオが安価に動画視聴を楽しむことができるサービスです。もし特に特定の作品を視聴したいというわけではなく、安価に動画配信サービスを使いたいというのであれば、プライムビデオ一択でしょう。
まずは無料体験

上記でご紹介した3つの動画配信サービスは、いずれも登録から一定期間は無料で全てのコンテンツを利用することができます。
登録後、もしその動画配信サービスが気に入らなければ、無料期間内に退会手続きを済ませれば利用料金はかかりません。
いくら料金が安くても見たい作品がなければ意味がない。いくら見たい作品があっても料金が高ければ躊躇してしまう……そんな不安も、無料期間を活用することで解消することができます。
TRY
FREE
他にも無料期間を設けている動画配信サービスは多数ございますので、一度試してみてご自分に合う動画配信サービスを見つけてください。
無料動画サイトでは駄目なのか

世の中には様々な動画サイトが存在します。YouTubeなんかは皆様もご存知でしょう。
他にもフランスの動画共有サービスであるDailymotionなどもありますが、それらの中には、ときに著作権を無視し違法にアップロードされた動画が存在します。
これらを視聴する行為は、その違法動画を使用している端末にダウンロードしない限り違法ではありません。インターネットを利用する行為そのものが厳密にはダウンロードしていることとほぼ同義ではありますが、この点は文化庁のウェブサイトにあります『違法ダウンロードの刑事罰化についてのQ&A』にも明記されています。
Q4:違法に配信されている音楽や映像を視聴するだけで、違法となるのでしょうか。
違法に配信されている音楽や映像を見たり聞いたりするだけでは、録音又は録画が伴いませんので、違法ではなく、刑罰の対象とはなりません。
違法となるのは、私的使用の目的であっても、著作権又は著作隣接権を侵害する自動公衆送信を受信して行うデジタル方式の録音又は録画を、自らその事実を知りながら行って著作権又は著作隣接権を侵害する行為です。
では、違法動画を視聴することの何が問題なのでしょうか。
ウィルスに感染

先述のYouTubeなどの有名動画配信サイトであるならば、単純に動画を視聴しただけではウィルスに感染する確率はほぼありません。
ですが、それら以外の違法動画配信サイトについてはその限りではありません。
表示される項目に、動画再生を開始するボタンに、あるいはサイトそのものに……どこにウィルスが仕込まれているか分からないのです。
もしご自身のパソコンがウィルスに感染した場合、
- データが勝手に削除される
- 操作が一切できなくなる
- 個人情報が抜き取られる
- 他の端末にウィルスをばら撒いてしまう
など、尋常ならざる害を被ることになります。
そして、データが削除されたり操作ができなくなる場合はともかく、感染したパソコンがウィルスをばら撒くことによって、最悪の場合、貴方自身が警察に逮捕される事態にもなりかねません。
実際にそういう事件がかつて起こったことを覚えている方もいらっしゃるのではないでしょうか。
パソコン遠隔操作事件(パソコンえんかくそうさじけん)とは、2012年(平成24年)の初夏から秋にかけて、日本において、犯人がインターネットの電子掲示板を介して、他者のパソコン(PC)を遠隔操作し、これを踏み台として襲撃や殺人などの犯罪予告を行ったサイバー犯罪事件。遠隔操作ウイルス事件(えんかくそうさウイルスじけん)とも呼ばれる。なお、報道等では、事件で使用された悪意のあるプログラムをコンピューターウイルスと表現しているが、正確にはトロイプログラムである。これら悪意のあるプログラム等は総称してマルウェアと呼ばれる。
最終的には誤認逮捕された方々は無事釈放されましたが、それでもできることならこんなことには誰も巻き込まれたくありません。
この事件で誤認逮捕された方々が違法動画サイトを利用していたわけではありません。
また、パソコンを使用していたら突然画面に『ウィルスに感染しています』などの表示が現れて驚いた経験をした方もいらっしゃるでしょう。そのまま、表示されたメッセージのとおりに操作を進めると警告は収まり一安心……ではありません。
これらの表示は偽のメッセージであり、悪意のあるソフトウェアをダウンロードさせることが目的です。メッセージに従ってそれらをダウンロードした場合、それ自体がウィルスか、危険なサイトへのアクセスを促すものである可能性が高いです。
スマホなら大丈夫?そんなことはありません

これらのコンピュータウィルスはパソコンのみをターゲットにしているからスマホやタブレットなら関係ない……なんてことはありません。
スマホに感染するウィルスはすでに無数に存在し、違法サイトを利用することでの感染のリスクはパソコンと同程度、あるいはそれ以上にあります。
殊スマホが爆発的に普及したことにより、インターネットに対する知識が不足したままネットを利用するユーザーが増加し、結果スマホでのウィルス被害はパソコンのそれよりも深刻化してしまう傾向にあります。
スマホだから大丈夫なんて理屈は、通用しないのです。
特殊詐欺被害

聞き慣れない言葉かもしれませんが、『オレオレ詐欺』『振り込め詐欺』などの総称です。これらのうち、違法動画サイトを利用していて遭遇する可能性が高いのは、『架空請求詐欺』でしょう。
違法サイトを利用したことによりメールアドレスなどが収集され、そのアドレスに対して架空請求のメールを送信し、振り込ませる……古典的ではありますが、これらは未だに被害が跡を絶ちません。
特に「自分は大丈夫だ」と油断している方が被害に遭いやすい傾向にあります。何事にも絶対に大丈夫というものはありませんので、十分に警戒するべきです。
映像業界全体の衰退

違法動画サイトで映像作品を見るということは、
アニメにしろドラマにしろ、映像作品を生み出すには莫大な費用がかかります。詳細は省きますが、その費用を取り戻す=利益を出すためにあるものが、作品の合間合間に差し込まれるCMであり、Blue-rayなどの映像メディアであり、主題歌CDなどの音楽メディアです。
違法動画サイトにアップロードされた動画には、CMはありません。アップロード者がその部分を削除しているからです。また、無料で見られるのならわざわざディスクを買おうとは思わないのが当然の流れでしょう。
そんな状況が続けば、当然制作する側はそこに利益が見出だせず、新たな作品を作り出すことはなくなります。
見たい作品、好きな作品を違法サイトで見続けることによって、見たい作品も好きな作品も消滅してしまうのです。
これを、『結局金か』とか『金を払いたいと思えるコンテンツを作れないのが悪い』というのは、そもそも間違っています。
動画配信サービスとレンタルを比較
冒頭にてすでに、レンタルショップでは見たい作品が貸出中だったり、そもそも置いていなかったりで借りることができないという状況に陥る可能性があります。
では、それ以外に動画配信サービスとレンタルとの違いはどの程度あるのでしょうか。
店舗レンタルとの違い

まずは、こちらが実際に店舗に訪れて作品をレンタルする場合と比較していきます。
さて、店舗でレンタルということを考える際、おそらく多くの方が『TSUTAYA』と『GEO』を思い浮かべるのではないでしょうか。
両者共にレンタルに対する料金は店舗ごとに若干差がありますので、全国一律この料金ということができません。そこでまずは、可能な限り同一のエリアにあるTSUTAYA、GEO両店舗の1枚あたりのレンタル料金を調査し、その結果からひとまず両者を比較してみましょう。
なお調査を行った店舗のレンタル料金は調査時点のものであり、今後変更される、あるいはすでに変更されている場合がございますのでご了承ください。
| 種類 | 日数 | TSUTAYA | GEO |
|---|---|---|---|
新作 |
1泊2日 | 380円 | 310円 |
| 2泊3日 | 470円 | 360円 | |
| 7泊8日 | 720円 | 650円 | |
| 旧作 | 7泊8日 | 360円 | 100円 |
| 延滞 | 200円 | 270円 | |
ご覧のように、当方の調査においては、TSUTAYAとGEOではGEOのほうが若干安いという結果になりました。
しかしながら、この時点で店舗レンタルよりも動画配信サービスのほうが手軽さもコスパも圧倒的に上であることは明らかでしょう。
仮に上記店舗のGEOで旧作を10本借りた場合、料金は単純計算で1000円ですが、その10タイトル以外に見ることはできず、また、レンタルしてから1週間後には返却しなければなりません。
これが、こちらも仮にhuluの場合、ほぼ同額で圧倒的多数の作品をいつでもどこでも好きなときに見ることができるのです。
しかも、他の誰かが借りていて自分がレンタルできないという事態も発生しません。
加えて、動画配信サービスなら、新作のみ料金が高くなるという状況もなく、常に一定の金額で新作も旧作も楽しめますから、そもそも比較のしようがありません。
更に店舗レンタルでは、借りるにしろ返すにしろ実際にそのお店に赴かなければなりませんが、動画配信サービスにはその手間が存在しません。
宅配レンタルとの違い

では、家からほとんど動くことなく動画作品のレンタルが可能な宅配レンタルはどうでしょうか。
宅配レンタルとは読んで時のごとく『自宅でレンタルの手続きを行い、後日レンタルした作品がポストに届く』方式のレンタル方法です。利用されたことがある方もいらっしゃるのではないでしょうか。
先述のTSUTAYAもGEOも宅配レンタルサービスを展開しており、それぞれに複数のプランが用意されています。今回はその中から、GEOの『スタンダード8』とTASUTAYAの『定額レンタル8』を見てみましょう。
| TSUTAYA | GEO | |
|---|---|---|
| 月額料金(税抜) | 1,865円 | 1,860円 |
| 月レンタル枚数 | 8枚 旧作借り放題 |
8枚 |
| 発送 | 2枚ずつ | 2枚ずつ |
料金そのものについては両者ともに大差はありません。ですがTSUTAYAの定額レンタル8は、1ヶ月の間に8枚レンタルし終えた後も、旧作や話題作を追加料金無しでレンタルし続けることができます。
つまり視聴可能作品数だけを見るなら、実質動画配信サービスと同等のサービスといえなくもありません。
ですがこの借り放題プランには新作や準新作は含まれておらず、また現物を配送している都合上どうしても手元に届くまでのタイムラグが発生します。
おおよそですが、レンタルの手続きを行って手元に届くまで2日~4日かかり、返却の際もまた同程度の日数がかかります。1ヶ月を30日と仮定すると、多くても14枚(2日で着荷→その日のうちに視聴して返却→2日で返却完了として計算)がレンタル上限でしょう。
動画配信サービスではその配送のタイムラグもなく、配信タイトルは新作旧作問わず何本でも見ることができます。料金的にも、などと大差はありませんし、サービスによっては宅配レンタルの半額以下で利用できます。
また、これは店舗レンタル、宅配レンタル双方にある問題ですが、ディスクに傷が入っていて映像や音声が乱れたり、そもそも視聴が不可能である場合もあります。動画配信サービスにはそのような心配がありません。
動画配信サービス一覧

正式な手続きを踏んで動画を配信している動画配信サービスなら、ウィルスに怯えることもなく、架空請求も届かず、業界が衰退することもありません。
見たい作品を探してレンタルショップをはしごする必要もなく、今まで知らなかった素晴らしい作品に出会う機会にも恵まれます。
それらがネット環境と端末さえあればいつでもどこでも楽しめるというのは、ネット普及の恩恵の1つといえるでしょう。
そんな数あるVODの中でも定額見放題を展開している動画配信サービスをピックアップしてご紹介いたします。
U-NEXT

株式会社U-NEXTが運営する、国内最大級の配信作品数を誇る動画配信サービスです。
その配信作品数は見放題のものだけでも80,000本以上、レンタル作品も含めると合計で130,000本以上もの作品がどこにいても楽しむことができます。
作品数も非常に魅力的ですが、何よりもそのタイトル追加の速さも特徴の1つ。例えば、日本では2018年8月1日に公開が開始された『インクレディブル・ファミリー』は同年の11月21日から配信が開始されていますし、2018年秋アニメもほぼ全ての作品が配信されています。
Amazonプライムビデオ
ネット通販最大手Amazonが運営する動画配信サービスです。プライムビデオ単独での会員登録ではなく、Amazonプライム会員としての登録が必要です。
見放題の配信作品数は公表されていませんが、月額換算400円という安さが非常に魅力です。
またそれに加えて、Amazonにある一部の楽曲がストリーミング可能になる『プライムミュージック』や注文した商品をいち早く届けてもらえる『お急ぎ便』の無料化など、Amazonプライム会員のサービスは勿論受けることができます。動画配信サービス以外にもよくネット通販を利用する方には、非常に優れたサービスといえるでしょう。
Hulu
HJホールディングス株式会社が運営する動画配信サービスです。
元はアメリカの動画配信サービスであり、海外ドラマに関してはトップクラスの配信数を誇ります。
更に、FOXチャンネルなどのストリーミング配信も行われており、『海外ドラマを見るならとりあえずHulu』といっても過言ではありません。勿論映画、国内ドラマ、アニメも充実しています。
Netflix

Netflix社運営の動画配信サービスです。
世界最大規模のユーザー数を誇るVODで、日本進出は後発ながらも着実にユーザー数を増やしています。
オリジナル作品を中心に展開されており、総配信数は他の動画配信サービスと比べると少なめです。その分質で勝負といった具合。また、視聴履歴によって個人個人に合わせたオススメタイトルを提示してくれるため、新しい作品に出会いやすい特徴があります。
dTV

NTTドコモが運営する動画配信サービスです。
かつてはドコモ契約者でなければ利用できませんでしたが、現在では誰でも登録が可能となっています。
アニメやドラマのみにとどまらず、ライブ動画やオリジナルコンテンツも豊富に用意されており、流石にプライムビデオほどではありませんがコスパは非常に高いです。ただ、単純にアニメを見たい場合は後述のdアニメストアやU-NEXT、海外ドラマならばhuluのほうが見られる作品数が段違いに多いです。
dアニメストア
その名の通りアニメに特化した動画配信サービスです。こちらもdTV同様に、現在ではドコモユーザーでなくても利用が可能です。
アニメ作品数では他を圧倒しており、その数は2,700作品以上。その内容も、旧作から最新作まで非常に多数を網羅しています。
その月額も400円(税抜)とコスパにも優れ、アニメだけを見たいならばプライムビデオ以上のサービスです。最新タイトルもほぼ網羅されているため、動画配信サービス利用の目的がアニメならほぼ一択ではないでしょうか。
auビデオパス

KDDIが運営する動画配信サービスです。
こちらもdTVやdアニメストア同様auユーザー以外は利用できませんでしたが、現在では他の携帯会社のユーザーでも登録が可能となっています。
料金も562円(税別)と比較的安価で、加えてTOHOシネマズなどの映画鑑賞券が割り引かれる特典などもついています。更に、毎月レンタル作品1本文のポイント(コイン)も付与されるため、コスパは比較的高いといえます。惜しむらくはdTV同様、他の動画配信サービスと比較するとやや配信タイトル数が弱い点でしょうか。
DAZN
イギリスのパフォーム・グループによって運営される動画配信サービスです。
他の動画配信サービスとは違いスポーツに特化しており、国内外の様々なスポーツ中継を視聴することが可能です。現状では、スポーツ中継を見るならこちらがマストアイテムです。
もし見たい中継に間に合わなかったとしても見逃し配信が用意されているため、国外のスポーツで時差的に視聴が厳しいものでも見ることができます。
動画配信サービスの[メリット][デメリット]
ここまで『違法動画サイトの危険性』や動画配信サービスそのものについてご紹介いたしました。
ここからは、改めて動画配信サービスにあるメリットとデメリットをそれぞれご紹介いたします。
動画配信サービスのメリット
まずは動画配信サービスを利用するメリットから触れたいと思います。
いつでもどこでも動画を楽しめる
これは動画配信サービスの最たるメリットではないでしょうか。
今やスマホさえあれば、よほどの僻地でもない限りどこにいてもインターネットに接続できる時代です。それはつまり、どこにいても動画配信サービスを利用できるということでもあります。
極端な話、電車などでの長い移動時間に動画を見るために事前にダウンロードしておく必要はもうないのです。
シリーズを一気に見ても金額が変わらない
レンタルですと、実店舗ならばディスク1枚に対し、ネット店舗ならば映像1つに対しレンタル料金がかかってきます。当然、作品の本数が増えれば増えるほどそのレンタル料も増加します。
ですが、定額見放題の動画配信サービスならばどれだけ長い作品を連続視聴してもかかってくる金額は変わりません。
配信の有無はともかくとして、ドラえもんを1話から最新話まで見ても、一定の金額で見ることができると考えると、そのコスパの高さは尋常ではありません。
レンタルにない作品も見られる
映像化された作品の全てがレンタルできるようになるわけではありません。
古い作品の場合、そもそもディスクメディア化していないことも少なくなく、以前はその際はプレミア価格のVHSやLPなどに手を出すか諦めるかしかありませんでした。
それが動画配信サービスの登場により、レンタルになかったりそもそも販売すらされていない作品であっても、手軽に見ることがd家いるようになったのです。
地方で放送されない作品も視聴できる
主にアニメをテレビ視聴する際の問題点ですが、全ての地域で全ての作品が放送されるわけではありません。
殊深夜アニメにおいてそれは顕著であり、関東や関西地方では放送されているのに九州や北海道では放送されていないタイトルはザラにあります。
以前ならそんな番組をリアルタイム視聴するには、それらが放送されている専門チャンネルがあるCATVやBS、CSに加入するか、それこそ違法にアップロードされた動画に頼るしかありませんでした。
それが動画配信サービスの台頭によって、作品次第ではテレビ放送よりも速いタイミングで視聴することが可能となりました。
勿論全てがテレビ放送と同じタイミングで配信されるというわけではありませんが、どうしても見たい作品が見られず、円盤を待たざるを得ない状況に陥ることがなくなった点は非常に高評価です。
動画配信サービスのデメリット
続きましてデメリット。一見万能の動画配信サービスにも、デメリットはあります。
通信量が大きい
自宅でパソコンから、あるいはWi-Fiをつないだスマホやタブレットからの視聴ならば通信量を気にする必要はありません。ですが通常の携帯端末用の回線から視聴を行う場合、どうしても通信量が大きくなり、通信制限がかかってしまうことがあります。
動画視聴の際の通信量は、動画が長ければ長いほど、画質がよければいいほど大きくなります。モバイル回線でも比較的安定して見ることができるHD画質の動画でも、1時間の視聴でおおよそ1GB弱の通信量がかかってしまいます。
事前に動画をダウンロードしておける動画配信サービスならば通信量を気にする必要はありませんが、突発的に何か見たくなったときには、どうしてもこの通信量はネックとなってくるでしょう。
作品によっては視聴期限がある
動画配信サービスでは実に多くの作品を視聴することができます。しかし、それら全てが未来永劫いつでも見られるというわけではありません。
人気が高く、安定して視聴者を獲得できているコンテンツならば長期間配信し続けることもありますが、反面、あまり人気が出なかったりするとすぐにライブラリから消去されてしまいます。
またそれ以前に、そもそも期間限定と銘打たれて配信されるタイトルもあります。見よう見ようと思って後回しにしていて、気づいたら配信が終わっていた……こうなってしまうと、再度配信されるのを待つか、映像メディアを購入するしかなくなります。
全てが1つのサービスで見られるわけではない
作品のラインナップは動画配信サービスによってまちまちです。作品Aと作品Bが見たいのに、配信サービスCには作品Aしかなく、配信サービスDには作品Bしかないことはままあることです。サービスの運営元にしてみれば、1人でも多くのユーザーに利用してもらいたいのですから、仕方のないことではありますが。
具体例を挙げるならば、2018年秋アニメの『色づく世界の明日から』はプライムビデオ限定配信で他の動画配信サービスでは視聴できず、しかしプライムビデオでは『ゴールデンカムイ』を見ることができません。
アニメに限らず、国内外のドラマや映画においてもこの状況は発生しています。
まとめ
動画配信サービスにもいくつかのデメリットは存在します。ですが、それを差し引いても受けられる恩恵が多彩であり、各サービス共に年々その利用者数を増やしています。
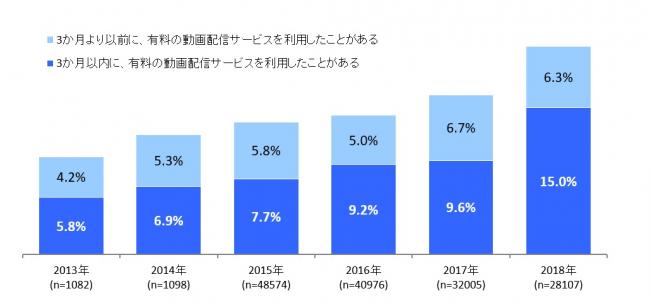
-有料の動画配信サービス利用率は15.0% | 株式会社インプレス
今後、動画配信サービスには更なる需要が集まることは間違いありません。
これを機会に、是非ご自身に合う動画配信サービスを見つけられてはいかがでしょうか。